|
|
|
|
|
Since temperature is among the most limiting factors in alpine environments and snow cover plays a key role in habitat characteristics and species distribution, changes in temperature and snow cover duration will have profound effects for alpine environments. For example, different communities exhibit varying susceptibility to these changes and within communities, there can be large differences among plant species in their responses. Other indirect effects of temperature, e.g. on soil moisture, wildfire frequency or insect outbreaks, may locally be very important and exceed direct effects of increased temperature or changes in precipitation. Species may respond by adaptation and/or migration to warmer temperatures, yet a prime concern for alpine ecosystems is that the rate of change will outpace the ability of species to adapt. There is a growing body of evidence that in the course of such rapid climatic changes genetic variation will be lost. |
|
|
|
Physiological responses and growth Environmental manipulation experiments showed that species respond specifically to each factor of environmental change. Plants generally respond most strongly to nutrient, particularly nitrogen, addition. |
|
Differences in growth responses due to increased temperature Most alpine plants respond to increases in temperature with increased vegetative growth as well as increases in reproductive effort (fig. 1), yet speed and amplitude of individual plant responses differ amongspecies (table 1) and sites (fig. 2). Warming experiments differ quite substantially in their results. This may partly be caused by differences in responses between different plant communities, but also by differences in experimental methodology: Warming by OTCs usually leads to increased air temperature within OTC, but partly also to increased soil temperature, reduced soil moisture and earlier snow melt. But there are also site-specific responses within the same species within single experiments. E.g. Carex stenantha var. taisetsuensis highly increased its canopy volume with the OTC treatment at site F2, while OTC effect was not detected at site F1 (for site description see fig 2). Probably site conditions have been severe for this species at the higher fellfield site, leading to a positive response to warming conditions, whereas at the lower site, growth conditions seem to be less stressful. Snowbed species usually show no positive response to temperature increase. In contrast, they typically are sensitive to spring frost, which may increase with higher temperatures and earlier snow melt. Nival species show (almost) no response until now. Species, which do not (or only slightly) respond to climatic warming (e.g. Ranunculus glacialis) may be particularly vulnerable to climatic change, especially if their habitat is invaded by other species with higher phenotypic plasticity and a better competitive ability. Many invertebrates also react with increased population growth to climatic warming, at least as long as desiccation is not induced. |
1 - Growth response to warming (127K)
2 - Growth responses of different alpine plant communities to warming (106K) |


|
Physiological responses e.g. changes in amount of secondary compounds Only few experiments investigated physiological responses of alpine plants to global change. Yet, it has been shown that increased N availability and temperature can affect the allocation to carbon-based secondary compounds which are important for plant resistance to biotic and abiotic environmental stresses such as herbivory, pathogens, or UV radiation. Again there are large differences between plant species. |
|
|
|
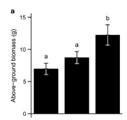
With increasing temperature and less amount of snow, the length of the growing season in temperate and northern alpine ecosystems is expected to increase in future. Phenological studies already showed a temperature driven extension of the growing period by up to two weeks in the second half of the 20th century, yet with regional differences. So far, earlier onset of the growing season has been observed in southern Scandinavia and in lower alpine regions in the Alps and other mountain regions, whereas in northern Scandinavia and at higher altitudes, such trends have not been observed yet. Whereas some species directly respond to the time of snowmelt, others are rather sensitive to the length of the photoperiod. Even if plants start to grow and/or flower earlier due to advanced snowmelt, they may not be able to effectively use this phenological shift for higher growth or fecundity rates: In a transplant experiment using Campanula thyrsoides, plants transplanted to the lowest site responded with a phenological shift, but at the same time, above-ground biomass, plant height, and number of flowers were reduced (fig. 3); so earlier onset of growth obviously caused a reduction in fitness in this species. |
3 - Responses of fitness-related phenotypic traits in Campanula thyrsoides |
|
Earlier snowmelt is often correlated with colder spring air temperatures and more frequent frost events experienced by above-ground plant parts resulting in enhanced frost or frost-desiccation damage (fig 4). Flower buds of some plant species like Delphinium barbeyi, Erigeron speciosus, and Helianthella quinquenervis are sensitive to frost, and the earlier beginning of the growing season in recent years has exposed them to more frequent mid-June frost kills. From 1992 to 1998, on average 36.1% of Helianthella buds were frosted, but for 1999-2006 the mean is 73.9%. For all three of these perennial species, there is a significant relationship between the date of snowmelt and the abundance of flowering that summer. Greater snowpack results in later snowmelt, later beginning of the growing season, and less frost mortality of buds. The loss of flowers and, therefore, seeds can reduce recruitment, and may also have consequences for pollinators, herbivores, and seed predators relying on these plants as food source. |
4 - Relationship between Helianthella quinquenervis flower heads not killed by frost and the first date of bare ground (119K) |

|
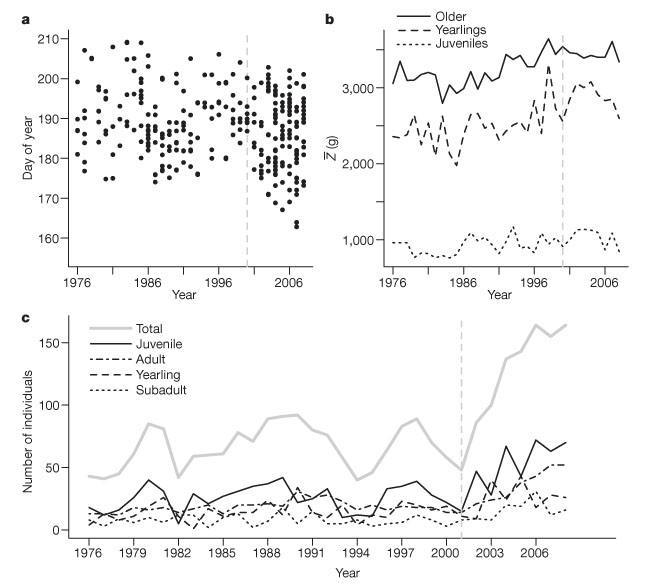
Responses of animals Some animals also respond to warmer spring temperatures, e.g. hatching of young ptarmigan became significantly earlier from 1975 to 1999. Marmots near the Rocky Mountain Biological Lab at Gothic now emerge about three weeks earlier than about 30 years ago. Earlier emergence from hibernation and earlier weaning of young has led to a longer growing season and larger body masses before hibernation, resulting in a decline in adult mortality. This in turn triggered an abrupt increase in population size since 2000 (fig 5). |
|
5 - Trends in phenology, mean phenotypic trait and demography for females of a yellow-bellied marmot (Marmota flaviventris) population in the Upper East River Valley, Colorado, USA. a Time of weaning (-0.17 days per year, P , 0.01), b mean 1 August mass (Z), c abundance in each age class (c). The four age classes are juvenile (< 1 yr), yearling (1 yr-old), sub-adult (2 yrs-old) and adult (>3 yrs-old). Sub-adult and adult masses are combined (older) in b. Vertical dotted lines delineate different phases of population dynamics (from Ozgul 2010). |
6 - Yellow bellied marmot (201K) |
;)
13 August 2018 |
||
| |
||